

-
神经生长因子启动哮喘神经.内分泌.免疫网络功能失衡
神经生长因子启动哮喘神经.内分泌.免疫网络功能失衡
【摘 要】 神经生长因子是一种对神经生长、分化起到营养作用的肽类,其在哮喘发病过程中被认为是连接神经.内分泌.免疫网络的桥梁,作用机制可能如下:a.神经生长因子引起气道神经解剖结构和功能变化,促进气道神经末梢合成和释放递质,有助于气道重构的形成;b.神经生长因子能够增强变应原特异性I g E抗体的表达,促进肥大细胞、嗜酸性粒细胞、淋巴细胞等在气道聚集,诱导其释放炎症介质,改变免疫应答平衡状态;C.神经生长因子可能启动肾上腺髓质细胞冗余性,使其向神经细胞转变,导致髓质细胞内分泌功能削弱,使肾上腺素合成、释放和再摄取功能障碍,最终导致循环中肾上腺素达不到维持气道舒张状态所需水平。
【关键词】 哮喘 神经生长因子 神经 内分泌 免疫网络
炎症性疾病,其发病机制与神经、内分泌和免疫系统密切相关.免疫细胞除了分泌神经递质和内分泌激素之外,其表面还广泛存在这些递质和激素的受体,这表明神经系统和内分泌系统产生的信号分子能通过受体调节免疫细胞的功能,反过来,免疫系统产生的信号分子亦可能通过类似的途径作用于神经内分泌系统,这种双向作用使多个系统内或系统之间得以相互交通和调节即构成神经一内分泌一免疫调节网络研究提示,神经生长因子(n e r v eg r o w t h f a c t o r。N G F)n-]O~是调节哮喘神经.内分泌.免疫网络失衡机制中的启动因素[2.--4].
1 N G F的生物学特性及其在哮喘中的表达情况
N G F是一种对神经生长、发育和分化起到重要作用的神经肽类,主要由神经细胞产生.免疫细胞(嗜酸性粒细胞、淋巴细胞、肥大细胞等)及组织特异性细胞(上皮细胞和成纤维细胞等)均能够分泌N G F,但在生理状态下,这些非神经细胞所分泌的N G F微乎其微,而在炎症状态下其分泌的N G F异常增加.N G F有两种受体:低亲和力受体p 7 5和高亲和力受体t r k A,t r k A是N G F的功能性受体,目前对p 7 5在N G F信号传导中的作用还不完全清楚,可能参与调节t r k A的活性.研究显示,靶源性N G F主要由靶组织释放,与感觉神经末梢上的N G F特异性受体结合后,逆行至感觉性节神经元的胞体,进而促使胞体内的感觉性神经肽表达上调研究表明,过敏性疾病特别是哮喘患者体内N G F水平显著高于对照人群,且其升高水平与患者病情严重程度呈正相关.B r a u n等发现,哮喘及其他变应性疾病发作时,气道内嗜酸性粒细胞、肥大细胞等均能合成和分泌N G F t g 9 1.研究显示,哮喘患者痰中巨噬细胞、淋巴细胞中N G F表达显著高于对照组,且在急性发作期升高更为明显。我们的研究表明,用N G F抗体对哮喘大鼠进行干预,在降低体内N G F表达的同时可以使气道炎症得以缓解。
2 N G F对哮喘发病机制中神经系统的影响
N G F能增加神经元的兴奋性,H u n t e r等[1 2]发现,向哮喘豚鼠气道内滴入N G F时,可以诱导豚鼠气道内含速激肽类的神经纤维数量明显增加。H o y l e等【l 3]发现,与对照小鼠相比,N G F转基因小鼠气道内含速激肽类感觉神经纤维数量显著增多,并且小鼠气道神经解剖结构和功能发生变化:神经兴奋性和递质合成增加、气道高反应性及气道平滑肌痉挛出现,从而促使气道重构形成。气道受复杂的自主神经支配,除经典的肾上腺素能、胆碱能神经外,还存在非肾上腺素能非胆碱能(n o n.a d r e n e r g i c n o n-c h o l i n e r g i c,N A N C)神经.N A N C神经分为非肾上腺素能抑制性神经和非胆碱能兴奋性神经,前者递质为血管活性肠肽(v a s o a c t i v e i n t e s t i n a l p e p t i d e,V l F),具有舒张支气管作用,而后者递质为P物质(s u b s t a n c e P,S P)等感觉神经肽,具有强烈收缩支气管作用.哮喘患者尸检发现其气道中s P阳性神经元明显增多[1 4】.有研究显示,哮喘豚鼠脊神经节中S P表达增加,并通过感觉神经末梢释放至呼吸道,从而增加血管渗透性、诱发支气管痉挛,参与哮喘的发生。我们也发现,与对照组相比,哮喘大鼠肺组织(主要为免疫炎症细胞和气道结构细胞)中N G F及其mR N A表达显著增加,其背根神经节中N G F蛋白表达增高,而N G F mR N A水平无明显变化,提示哮喘状态下N G F增高可能是由于支气管肺组织合成N G F增加所致,背根节可能是N G F储存及发挥效应的场所.同时还发现,与正常大鼠相比,哮喘大鼠肺组织中S P及其mR N A表达均无明显变化,而其背根节中S P及其mR N A表达显著增高,这可能与速激肽在效应器官中的不稳定性有关产生效应后立即被灭活.用N G F抗体对哮喘大鼠进行预处理后,则其背根节细胞中S P及其m R NA的表达则显著下调。上述资料提示,哮喘发病过程中可能存在链式效应:气道上皮、炎症细胞(嗜酸性粒细胞等)合成N G F增加,合成的N G F被转运到脊髓背根节中储存,并刺激神经节合成S P增多,S P再通过神经突触分泌,作用于气道效应细胞(上皮细胞、平滑肌细胞等而触发气道神经源性炎症,反过来,炎症反应又可以上调肺组织中N G F的表达,形成恶性循环。
3 、N G F对哮喘发病机制中免疫系统的影响
研究证实,N G F能增加大鼠肥大细胞数量,刺激肥大细胞脱颗粒,此外,N G F可刺激人外周血嗜酸性粒细胞释放炎性介质[1刀.有研究提示,人C D 4 T细胞能够克隆表达t r k A,并能产生和释放N G F【l 8】,B淋巴细胞上亦有N G F功能性受体存在,并且发现t r k A与B淋巴细胞早期信号转导机制有关嘲。
哮喘患者在吸入变应原后出现变态反应,表现为气道平滑肌收缩、黏液分泌和血浆渗出增加.其中变应原与I g E抗体结合后能够促使肥大细胞释放组胺等炎性介质.N G F能够增强变应原特异性I g E抗体的表达,促进肥大细胞增殖、分化及释放炎症介质,d e V r i e s等发现,N G F抗体干预能够缓解变应原诱发的急性支气管收缩.从而提示放大变应原诱发的速发型变态反应可能是N G F在哮喘发病过程中的作用机制之一.在气道变应性炎症中,p 7 5受体可介导嗜酸性粒细胞向炎症组织中募集,p 7 5受体缺失时,变应原刺激则不能增加气道中N G F的表达、嗜酸性粒细胞浸润和炎症介质几.4、几.5等的释放冽。N a s s e n s t e i n等例研究表明,变应原刺激后,哮喘患者气道嗜酸性粒细胞中p 7 5表达增加.哮喘患者嗜酸性粒细胞与NG F共同培养后能够使细胞的存活能力增强、活性增加,从而提示N G F能够增强变应原激发的气道嗜酸性粒细胞炎症.这些资料提示,N G F可能通过诱导嗜酸性粒细胞、刺激T h 2细胞因子等作用达到激活炎症网络的病理生理效应.
4 N G F对哮喘发病机制中内分泌系统的影响
气道平滑肌缺乏肾上腺素能神经支配,其气道舒缩调节主要通过循环中肾上腺素作用于气道平滑肌肾上腺素能受体得以实现.肾上腺素由肾上腺髓质合成、释放入血,能拮抗各种刺激引起的支气管收缩.通常,当机体受到不利刺激或出现“紧急”状态时,交感.肾上腺髓质系统释放大量肾上腺素来应对刺激带来的负面效应.哮喘发作无疑是一种紧急状态,然而研究表明,哮喘发作患者循环中肾上腺素水平与对照者相比并无明显升高,同时发现,足以诱发哮喘的运动不能引起哮喘患者肾上腺素水平发生显著变化,但等量的运动却能使对照者肾上腺素水平显著升高捌.由此可见,哮喘患者可能存在肾上腺素释放障碍。
肾上腺髓质细胞在分化障碍状态下可出现肾上腺素合成、释放减少,但其具体发生机制不清踟.“细胞冗余性”(c e l l u l a r r e d u n d a n c y)概念的引入为细冯俊涛等:神经生长因子启动哮喘神经.内分泌.免疫网络功能失衡·2 4 3。胞分化障碍所致肾上腺髓质内分泌功能改变提供了理论依据:冗余性是指除必须具备的功能外,还具有潜在的“多于必需”的能力.简言之,即一种细胞除具有固定的“专职”功能外,还具有潜在的“兼职”功能.在疾病、人为干预等条件下,细胞冗余性启动,导致细胞“专职”与“兼职”功能失调.肾上腺髓质在胚胎早期和交感神经并行发育,之后,侵入原始肾上腺皮质,发育为髓质而逐渐产生内分泌功能.由于肾上腺髓质细胞与神经细胞在发育过程中具有同源性,使得这种细胞除内分泌功能之外,还具有潜在的神经功能(向神经细胞转分化能力1.研究发现,N G F对正常肾上腺髓质嗜铬细胞干预后,这些细胞能够长出突触,在细胞形态、生化及生理等性状方面向交感神经元转化.交感神经合成、分泌去甲肾上腺素和多巴胺等,但不产生肾上腺素,那么,哮喘发作过程中肾上腺素释放障碍是否系N G F或其拟似物启动肾上腺髓质细胞冗余性使其向神经细胞转变所致?
我们研究发现:与对照组相比,哮喘大鼠体内肾上腺素水平不仅没有升高,反而下降,同时,哮喘大鼠体内肾上腺素限速酶P N MT的表达亦显著降低,电镜下哮喘大鼠肾上腺髓质细胞出现细胞间隙增宽、线粒体增多、嗜铬颗粒减少,其细胞膜可以看见杵状和绒毛状突起,近细胞膜处有囊泡形成划.这些结果表明,N G F可能通过启动肾上腺髓质细胞的冗余性使其表型和功能向神经细胞转变,导致肾上腺素水平下降而参与哮喘的发生.
哮喘患者体内N G F水平显著高于正常人,且与病情严重程度呈正相关,随后发现N G F转基因小鼠对卵白蛋白诱发哮喘的敏感性显著高于野生型小鼠邳”.我们的前期研究也表明:用N G F抗体对哮喘大鼠进行干预能够减轻其气道炎症,并能减少肺组织及脊髓背根神经节中N G F和神经肽类物质的表达。
N G F由靶细胞产生后被神经轴突末稍摄取逆行转运至胞体,对神经递质传递及突触可塑性的调节有重要作用,N G F增高能够使S P等神经递质合成、释放增加.作为哮喘气道神经源性炎症发生过程中重要的神经肽,S P对肾上腺素的储存、释放和摄取也起到重要的调节作用.含S P的神经纤维在正常肾上腺皮质及髓质均有分布,逆行示踪法显示这些神经纤维来自脊髓背根神经节嗍.Z h o u等采用不同浓度的S P对大鼠肾上腺进行灌注,发现低浓度的S P能够促进烟碱及电刺激作用下肾上腺素释放,而较高浓度的S P则抑制肾上腺素分泌.哮喘中气道结构细胞及免疫炎症细胞等均可产生N G F,N G F被神经轴突末稍摄取逆行转运到背根节,从而使背根节合成、释放S P增加.S P能够导致气道平滑肌收缩、粘液分泌、微血管渗漏、炎症介质释放以及气道高反应性发生。推测N G F增高可能导致哮喘肾上腺素释
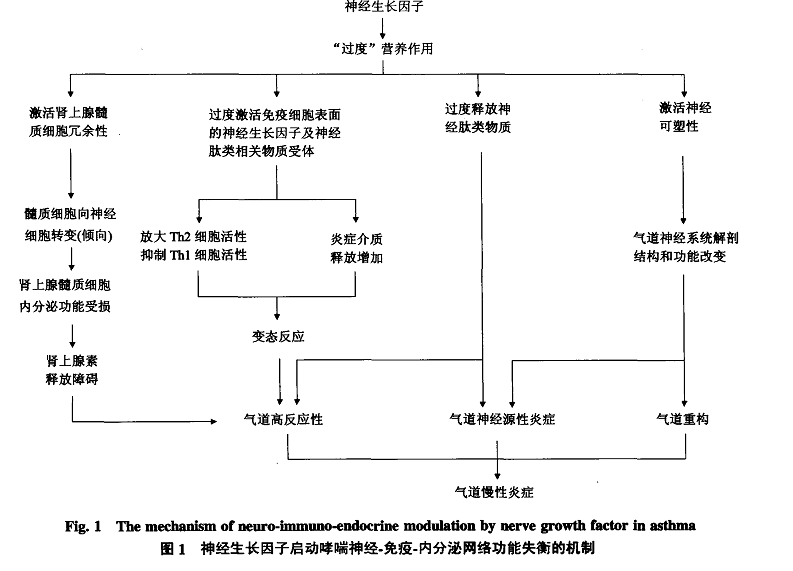
放障碍,具体机制可能为:a.N G F启动肾上腺髓质细胞冗余性,使髓质细胞向神经元转变,导致其内分泌功能削弱.b.N G F通过脊髓背根节及其支配的肾上腺髓质神经纤维来调控S P等递质,进而对肾上腺髓质细胞的生理、生化功能产生影响,使肾上腺素的合成、释放和再摄取出现障碍,最终导致循环中肾上腺素达不到维持气道舒张状态所需要的水平。
归纳起来,N G F在哮喘发病过程中的作用机制可能如下:在炎症环境中,靶细胞合成N G F增加,一方面,N G F能够直接作用于机体免疫细胞,造成免疫功能紊乱,另一方面,N G F被神经突触转运到脊髓背根节细胞中储存,并刺激神经节细胞合成S P等神经肽类物质增多,S P再通过神经突触分泌,作用于气道效应细胞(上皮细胞、平滑肌细胞等)触发气道神经源性炎症反应,反过来炎症反应又可以上调机体N G F的表达,从而形成恶性循环.除此之外,N G F可以通过前述途径使肾上腺髓质细胞向神经元转变,导致其分泌肾上腺素减少,诱导哮喘发作(图1).
.jpg)




