

-
妊娠期亚临床甲减大鼠对子代学习记忆能力及脑发育相关基因表达的影响
【摘要】目的通过建立妊娠期亚临床甲状腺功能减退(甲减)孕鼠动物模型,观察母鼠亚临床甲减对仔鼠行为学改变的影响,并从脑发育相关基因表达方面探讨母体亚临床甲减导致胎儿脑发育障碍的机制。方法将6 0只S P F级雌性成年Wi s t a r大鼠随机分成3组(每组n=2 0):(1)亚临床甲减组:手术切除甲状腺,于背部皮下植入微渗泵,L—T装人泵中持续泵入;(2)甲减组:手术完全切除甲状腺,术后注入等量生理盐水,作为阳性对照;(3)正常对照组(假手术组):行甲状腺切除假手术,术后注入等量生理盐水。取仔鼠出生后第3、7及2 1天海马组织,以实时定量P C R法检测各时间点脑源性神经营养因子(B D N F)、神经细胞黏附分子(N C A M)m R N A表达;以We s t e r n印迹法检测B D N F、R a p l的蛋白含量;第4 O天仔鼠进行M o r r i s水迷宫学习与记忆测试。结果(1)与正常对照组比较,亚临床甲减组的仔鼠第3天B D N F m R N A和蛋白表达减弱(均P
【关键词】亚临床甲状腺功能减退症 妊娠 脑发育
起的母体和胎儿甲减可以导致儿童的智力缺陷(呆小症、克汀病)早已被认识,母体亚临床甲减对儿童智力的影响近几年见于几项临床病例研究的报告。,并由此受到国内外内分泌学、围产医学和优生学界的广泛重视。目前国际上有关妊娠期母体亚临床甲减对后代脑发育影响的基础研究还鲜见报道。本研究旨在利用手术切除甲状腺、再植入左旋甲状腺素(L—T)微渗泵建立亚临床甲减Wi s t a r孕鼠模型,探讨母鼠孕期亚临床甲减对仔鼠学习记忆能力及海马相关基因表达的影响
材料和方法
一、材料
1.主要试剂和药品:L—T(S i g m a,美国),微渗泵(O s m o t i c Mi n i p u m p)A L Z E T~,型号2 0 0 1和2 0 0 2(A l z a,美国),T r i z o l试剂盒(I n v i t r o g e n,美国),S Y B RP r e m i x E x T a q T M(T a K a R a,日本),P C R引物由T a K a R a公司设计合成,兔抗脑源性神经营养因子(b r a i n—d e r i v e d n e u r o t r o p h i c f a c t o r,B D N F,C h e m i c o n,美国),兔抗R a p l(C e l l S i g n a l,美国),小鼠抗大鼠神经细胞黏附分子(n e u r a l c e l l a d h e s i o n m o l e c u l e,N C A M,a b e a m,美国),B C A蛋白定量试剂盒(P i e r c e,美国),内参照B.a c t i n、辣根过氧化物酶(H R P)羊抗兔I g G、羊抗鼠I g G(J a c k s o n,美国),E C L发光试剂盒及P V D F膜(A m e r s h a m,美国),其它试剂均购自S i g m a公司。
2.动物分组及亚临床甲减母鼠模型的建立:6 0只S P F级雌性Wi s t a r大鼠(购自北京维通利华实验动物有限责任公司),体重2 0 0~2 2 0 g,适应性饲养7 d后随机分成亚临床甲减组(n=2 0),甲减组(n=2 0)及正常对照组(假手术组;n=2 0)。1 0%水合氯醛(3 m l/k g)腹腔注射麻醉,参考E s c o b a r—M o r r e a l e等的方法制作亚临床甲减动物模型。手术完全切除甲状腺,术后恢复1个月之后,当雌鼠体重稳定在2 2 0 g左右时,将微渗泵(O s m o t i c M i n i p u m p)植入甲减鼠背部皮下,L—T装入泵中,以每天1.0 1 0 0 g体重浓度持续泵入。当血清T S H高于正常、而r I T r水平在正常范围时,提示亚临床甲减鼠建模成功。1 0 d后与正常的雄鼠合笼交配,雌:雄=2:l,当雌鼠阴道涂片镜下发现精子确定为妊娠0天,记为E 0。自E l 5始L—T剂量调整为每天1.0 5 1 0 0 g体重,持续输注至仔鼠生后1 0 d,将分娩当日定为P 0。甲减组:手术完全切除甲状腺。输注等量生理盐水,作为阳性对照。假手术组:仅手术暴露甲状腺,不切除甲状腺,术后输注等量生理盐水。为防止由于甲状旁腺的摘除而导致的低钙抽搐,术后饮水中均加入1.5%的葡萄糖酸钙。
二、方法
1.血清T S H、T T的测定:分别于母鼠E l 0、1 3、1 7,P 1(每日龄各1 5只)眶后静脉丛取血,分离血清,应用美国D P C公司固相化学发光酶免疫分析试剂盒测定血清T S H、1 T r。测定的灵敏度为1 2.8 7 n m o l/L。T S H批内C V 1.2 3%~3.3 8%,}比问C V 1.5 7%~4.9 3%,T Y批内C V 1.2 6%~3.2 0%,批间C V3.5 8%~6.6 7%。
2.动物灌注及标本收集:出生后仔鼠3、7及2 1 d断头处死,冰上迅速剥离两侧海马入匀浆器中,加入适量4℃预冷的组织裂解液,匀浆后收集于离心管4 o【=,1 2 0 0 0 g,离心1 5 r a i n后收集上清,分装后冻存于一7 0 c c。
3.实时定量P C R检测B D N F、N C A M m R N A水平:采用S Y B R G r e e n荧光实时定量P C R方法检测,以G A P D H作为参照,B D N F:上游引物5一A T C C A C T G A G—C A A A G C C G A A C一3,下游弓l物5一C A G C C T T C A T G C A A—C C G A A G T A.3;N C A M:上游引物5一A C A G C G G T G A A—C C G T A T T G G A一3,下游引物5一A A T r G T A C C T G T G C T—G T G C T G G A G一3;G A P D H:上游引物5一G G C A C A G T C—A A G G C T G A G A A T G一3,下游引物5一A T G G T G G T G A A—G A C G C C A G T A一3。取冻存海马组织约1 0 0 m g剪碎,用T r i z o l试剂提取总R N A,取4 g总R N A用随机引物和M—M L V逆转录成1 0 l c D N A,取2 l c D N A加用S Y B R P r e m i x E x T a q T M在A B I 7 5 0 0型荧光定量P C R仪上进行P C R扩增并检测荧光。反应条件:9 5 c【=预变性1 r a i n,9 4℃1 5 S,5 9 o C 3 0 S,7 2 o C 3 5 S,4 0个循环。计算每个反应管内的荧光信号达到设定阈值时所用的循环数(C t值),以G A P D H校正每个样品c t值,每个目的基因的c t平均值减去对应模板的内参基因(G A P D H)的c t平均值,得到A C t值,A A C t:A C t实验组一A C t对照组,则每个目的基因在某个模板中的平均相对含量为2“。
4.B D N F、R a p l蛋白表达检测(W e s t e r n印迹法):1 0 0 m g海马组织匀浆后提取蛋白、定量,取6 0 g的蛋白样品,加十二烷基硫酸钠(S D S)上样缓冲液,加热、离心后上样于非连续梯度S D S一聚丙烯酰胺凝胶上电泳,经转膜、封闭,加入相应蛋白抗体,二抗标记,增强化学发光法(E C L)显色,显影定影。凝胶成像系统中扫描,分析,计算各个条带的灰度值,以内参照B—a c t i n灰度值比值表示蛋白相对表达量。
5.M o r r i s水迷宫(M WM)行为测试:仔鼠出生后第4 0天进行M o r r i s水迷宫行为学检测,M o r r i s水迷宫D M S-2系统由上海吉量软件科技公司提供。(1)定位航行实验:实验历时5 d。实验前大鼠自由游泳1 d。正式实验每天训练4次,每次9 0 S,随机从东、西、南、北4个人水点选择一个,将大鼠面向池壁放入水中,记录大鼠寻找并爬上平台所需时间即逃避潜伏期(E s c a p e l a t e n c y)。若大鼠在9 0 S内未找到平台,由实验者将其引导至平台,逃避潜伏期记为9 0 S;大鼠在平台上停留1 5 S,然后从平台上取下休息3 0~6 0 S,再进行下一次训练。每只动物共计训练l 6次。计算每天各组4次逃避潜伏期的平均值。(2)空问探索实验:于第4 4天撤除平台,然后将大鼠随机从不同象限(除靶象限外即原平台所在象限)面壁置入池内3次,测量9 0 S内动物在靶象限和其他各象限的游泳时间,长时记忆能力测试于第4 5~4 9天进行,实验方法同第4 1~4 4天,安全台于第4 8天时撤除,在距离大鼠最后一次游泳时间2 4 h以后,于第4 9天测量9 0 S内动物在靶象限的游泳时间。
三、统计学处理
计数资料均以±s表示。数据采用S P S S 1 2.0统计学软件进行方差分析,组间比较采用5一Ⅳ一K法。以P
结果
一、孕鼠血清T S H、T T水平比较
与对照组比较,亚临床甲减组表现为血清T T水平未见明显改变,而T S H水平明显升高(P<0.0 5),符合亚临床甲减激素血清学改变;与甲减组比较,1丫r水平明显高于甲减组,T S H水平明显低于甲减组(P<0.0 1),提示亚临床甲减孕鼠模型建立成功(表1)。P 4 0进行M o r r i s水迷宫测试时,亚临床甲减组仔鼠血清激素水平和对照组无统计学差异(P>O.0 5)。甲减组仔鼠体重仍低,血清.r r水平接近试剂盒检测下限(1 g/d 1),显著低于同日龄对照组,T S H较同日龄对照明显升高(P
二、B D N F、N C A M m R N A表达量的变化(图1、图2)
与正常对照组比较,亚临床甲减组仔鼠在出生后3 d海马B D N F m R N A表达量下降2 7%(P<0.0 5),出生后7、2 1 d海马B D N F m R N A表达量与正常对照组间无显著差异(P>O.0 5);N C A M m R N A表达量有升高的趋势,但差异无显著性(P>0.0 5)。与亚临床甲减组和对照组比较,甲减组仔鼠B D N F m R N A表达量3、7和2 1 d均明显降低(P
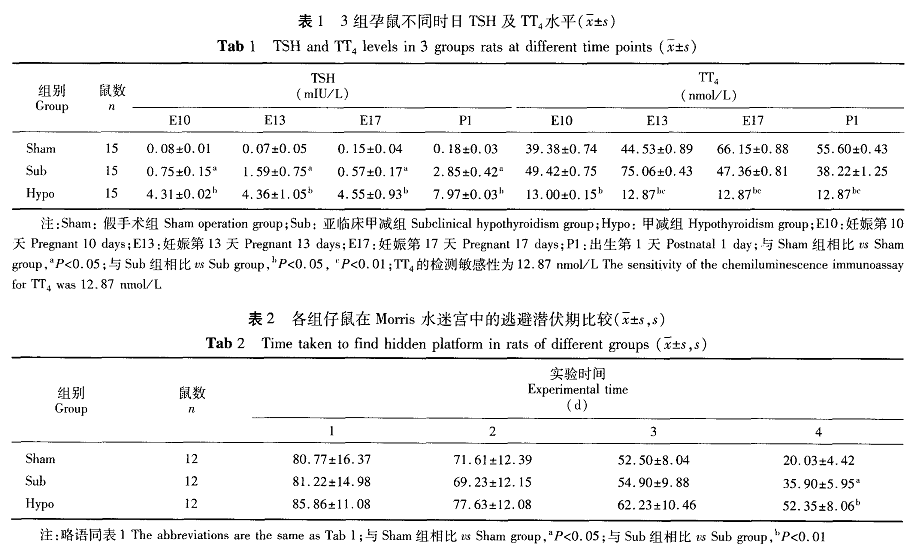
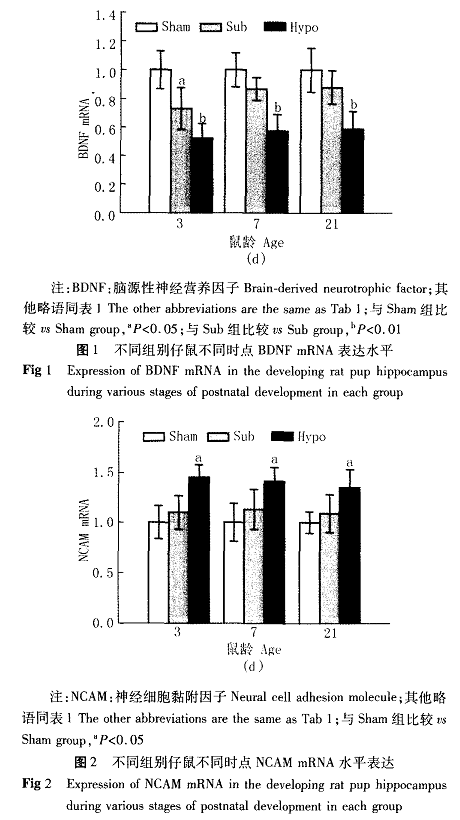
三、B D N F、R a p l蛋白表达量的变化(图3、图4)与正常对照组比较,亚临床甲减组仔鼠海马B D N F蛋白表达量在出生后3、7 d时明显降低,降低幅度分别为3 3%、2 9%(P
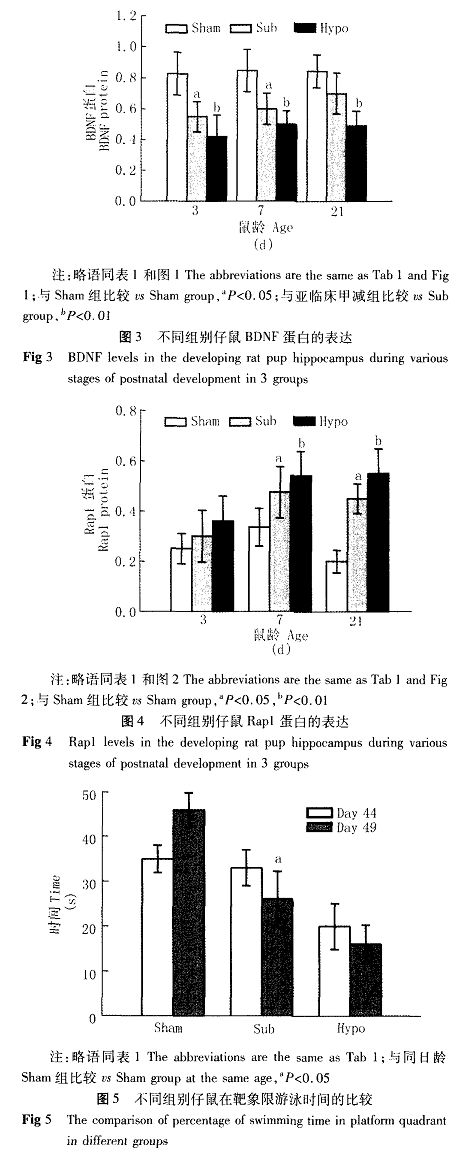
四、学习记忆行为学检测结果(表2、图5)定位航行实验的第1天各组大鼠均在适应期,故各组大鼠平均逃避潜伏期变化不明显,从训练的第2天始,各组仔鼠的平均逃避潜伏期随着训练次数增加而逐渐缩短,但差异无统计学意义(P>0.0 5)。实验第4天,对照组训练平均逃避潜伏期由(5 2.1 4±5.9 3)S迅速下降至(1 8.2 3±4.2 7)S,亚临床甲减组平均逃避潜伏期由(5 4.3 5±5.0 2)S下降至(3 5.6 3±5.9 5)S,甲减组平均逃避潜伏期(6 2.2 4±6.0 7)S缓慢下降至(5 5.1 6_+5.1 5)S,重复测量方差分析显示差异有统计学意义(P<0.0 5)。在空间探索测量仔鼠的长时空间记忆能力实验中(第4 9天),与正常对照组比较,亚临床甲减组仔鼠在靶象限活动时间显著降低,但明显高于甲减组(P<0.0 5)。
讨论
在胎儿自身下丘脑一垂体一甲状腺轴系统未完善之前(主要是妊娠前半期),对于神经系统发育,母体来源的T起着至关重要的作用。母体正常水平的T可以保护胎儿大脑避免T缺乏J。亚临床甲减的母亲,即便有正常水平的T,但升高的T S H已经提示甲状腺的储备功能下降。而妊娠期间由于母体对于甲状腺激素的需求量增加从而使甲状腺的工作负荷增大,所以并不能排除胎儿大脑缺乏T,从而影响胎儿脑发育和智力水平。因此,妊娠期母体亚临床甲减对胎儿第一阶段脑发育的影响成为近年来内分泌等领域的研究热点。传统的甲减动物模型主要采用甲巯咪唑或碘缺乏的方法建立,由于在造成母鼠甲减的同时都有可能影响胚胎发育,所以本实验采用手术切除甲状腺、再植入L—T微渗泵建立亚临床甲减母鼠模型。模型组血清检测提示T S H水平明显高于同期对照组,而T T水平仍波动在正常范围内,符合亚临床甲减激素血清学改变特点。
研究证实,在鼠脑发育期海马是接受甲状腺激素调控的敏感脑区之一。B D N F是甲状腺激素调节脑发育的重要靶分子之一,其在海马表达极为丰富。B D N F通过与神经元轴突末梢膜上的高亲合力受体酪氨酸蛋白激酶B(t y r o s i n e k i n a s e B,T r k B)结合,启动细胞内信号转导途径如R a s一丝裂原活化的蛋白激酶(M A P K)途径、磷酸肌醇3激酶(P I一3 K)途径等,参与细胞的分化、黏附、增殖与成熟等重要的生物学过程,。大多数情况下R a s—M A P K途径被认为是B D N F发挥作用的优势途径。B D N F可能直接参与神经递质释放、突触的可塑性重建,参与学习记忆的巩固等过程。突触长时程增强效(1 o n gt e r m p o t e n t i a t i o n,L T P)首先在海马发现,这是突触传递可塑性的证据,并已成为研究学习记忆过程的突触模式。记忆的获得和巩固与B D N Fm R N A表达的增加及其受体的活化有关,B D N F基因敲除小鼠,海马神经元L T P严重缺陷,给予外源性B D N F或转染B D N F基因,受损的海马L T P完全恢复。L i n n a r s s o n等¨应用M o r r i s水迷宫检测B D N F基因缺失突变小鼠的空间学习能力,结果发现青年B D N F突变小鼠空间学习能力明显下降,需要用对照组2倍的时间去完成任务。
C i r u l l i等通过研究评价了大鼠单次海马内注入B D N F对记忆保持的影响,B D N F处理组在M o r r i s水迷宫钡0试中表现出逃避潜伏期缩短,到达平台的路径短。提示B D N F基因表达在加强学习记忆过程和L T P中起着重要的作用。本实验结果显示,亚临床甲减仔鼠在第3天海马B D N Fm R N A明显下调,而B D N F蛋白表达量在第3天、7天明显降低。提示母体亚临床甲减早期即可通过调控B D N F基因表达影响仔鼠的脑发育,而B D N F蛋白的改变比m R N A水平的改变滞后。
R a s相关蛋白一1(又称K r e v一1或R a p 1)是一类小分子G T P结合蛋白,属于R a s家族成员。R a p l蛋白与R a s蛋白有5 0%的氨基酸相同,两者的分子结构域极其相似,R a p l通过结合与R a s相同或相似的效应分子而发挥完全不同于R a s的信号转导作用¨。目前已明确R a s的主要下游效应分子有R a f l、R a l G E F s和P I一3 K。R a p l通过捕获R a s下游的效应蛋白,尤其是R a f t,形成没有活性的复合物而干扰R a s蛋白发挥功能,从而影响R a s在神经细胞分化、增殖、神经递质传递等方面的生理作用¨。越来越多的研究证实R a s系统参与了海马L T P过程和学习记忆的形成,激活R a s依赖性的M A P K/细胞外信号调节激酶(E R K)信号途径是形成长期记忆所必需的¨。K e l l y等¨将R a s/E R K特异性抑制剂U O 1 2 6注入S D大鼠脑室,利用行为学方法检测后发现,与对照组比较,在第2天,实验组大鼠寻找新、旧目标所用时间均显著长于对照组,表明大鼠长时记忆保持和学习功能受到明显影响。
本研究证实,母体亚临床甲减导致了发育期仔鼠海马R a p l蛋白的表达升高。在正常对照组,R a p l蛋白在出生后第7天达到高峰,第2 1天,其表达水平已经降到约为第3天的一半。而在亚临床甲减组仔鼠海马的R a p l蛋白在第2 1天时仍处于高表达状态,甲减组这一现象表现得更为明显。业已证实,B D N F需要通过与受体结合激活R a s/E R K信号通路来发挥其对神经元的保护作用,本研究中,母体亚临床甲减时R a p l蛋白表达的增强,可能会拮抗R a s的作用,从而在信号传递的下游抑制B D N F促神经元发育和存活的功能。因此,母体亚临床甲减对发育期鼠脑引起的学习记忆和认知能力的障碍,可能是改变了海马B D N F的表达和抑制了B D N F—R a s信号转导通路的双重作用所致。在神经发育、再生以及与学习记忆巩固相关的突触可塑性等功能中,N C A M起着关键性的作用。N C A M中含有甲状腺激素的受体结合位点,直接受甲状腺激素调控。N C A M的正常表达是海马神经元存活、生长、出芽、突起及延伸的必要条件,在学习和记忆过程中发挥重要作用¨。在国外相关研究中,I g l e s i a s等¨用原位杂交方法发现甲减1 5日龄和2 0日龄的仔鼠海马N C A M基因表达量有增加的趋势,但其峰值始终低于对照组,他们认为,这是由于甲状腺激素缺乏延迟了N C A M正常的表达时间,导致N C A M表达时程发生了改变。但N C A M表达延迟的具体机制尚需进~步研究。本研究中,亚临床甲减组仔鼠N C A M表达量与对照组差异无统计学意义,可能是因为经过一段时间的代偿,N C A M的表达量趋于正常,其确切机制尚需进一步研究。
综上所述,妊娠期母体亚临床甲减能引起仔代出生后早期第3天海马B D N F m R N A表达下降,第3天、第7天B D N F蛋白水平表达下降。第7天和第2 1天海马R a p l蛋白表达水平的增加,R a p l为B D N F下游信号分子,表明妊娠期母体亚临床甲减的仔代学习记忆功能的损伤可能是二者共同参与的结果。
.jpg)




